Ультраструктура
Разработка методов электронной микроскопии вызвала революцию в изучении пластид, в том числе и хромопластов. Оказалось, что хромопластам присущи все характерные признаки пластид, но по ультраструктуре они существенно отличаются от хлоро-, лейко-, этиопластов и пропластид. Одновременно были показаны различия между отдельными хромопластами не только по форме, но и строению.
Применение электронно-микроскопических методов позволило выяснить процессы, происходящие при развитии хромопластов, взаимосвязь последних с другими типами пластид и идентифицировать в очищенном виде отдельные субструктуры, изучить их биохимический состав, а также некоторые аспекты функционирования.
Оболочка хромопластов
Подобно другим типам пластид хромопласты окружены оболочкой, или перистромиумом (в работах, опубликованных в последние годы, преимущественно используется термин "оболочка"). Некоторые сложности при обнаружении оболочки встречаются в исследованиях кристалловидных хромопластов. Светооптическая микроскопия при обычных условиях не позволяет разграничить оболочку от выкристаллизованного пигмента и стромы.
Однако при использовании изотонических растворов для фиксации оболочка отслаивается и выявляется в виде тонкого хрупкого образования, окружающего пластиду, которое растягивается по мере роста кристаллов. Исследования с применением электронной микроскопии показали, что оболочка хромопластов, как и оболочка других типов пластид, состоит из двух мембран.
В некоторых работах идентификация двух мембран, в особенности внутренней, затруднена. Б.Т.Матиенко и Е.М.Чебану считают, что недостаточная четкость контура внутренней мембраны оболочки может быть обусловлена плохой сохранностью материала во время подготовки к микроскопированию, хотя возможно, что она отражает естественное состояние деструкции или связана с плоскостью среза.
Электронно-микроскопические исследования подтвердили результаты светооптических наблюдений с использованием изотонических растворов о наличии мембран угкристалловидных хромопластов. Так, у хромопластов мякоти томатов, в которых пигмент (ликопин) выкристаллизовывается в виде малиновых игловидных или призматических образований, хорошо видна двух-мембранная оболочка, которую также наблюдали у игловидных клеток мякоти арбуза.
На электронных микрофотографиях кристалловидных пластид корнеплодов моркови две мембраны оболочки не всегда удается разграничить. Однако это не помешало А.Frey-Wyssling, F.Schweger его заключить, что двух-мембранная оболочка сохраняется на протяжении всего развития хромопластов корнеплодов оранжевых сортов моркови, в том числе и в период интенсивного роста игловидных кристаллов. 
При этом наружная мембрана хлоропластной оболочки проницаема для многих веществ и поэтому не может рассматриваться как эффективное регулирующее устройство. Однако внутренняя мембрана строго селективна в отношении проницаемости и свободно пропускает лишь незначительное количество самых простых молекул, таких, как CO2, O2; несколько затруднено проникновение гликолевой, фосфогликолевой, дикарбоновых и аминодикарбоновых кислот.
Незначительной проницаемостью обладают пентозо- и гексозофосфаты. Не проникают через внутреннюю мембрану хромопластов НАД и НАДФ, пентозы, гексозы, нейтральные аминокислоты, би- и полифосфаты (включая бифосфаты Сахаров и неорганический пирофосфат), мевалоновая, уксусная и другие кислоты. В оболочке хлоропластов имеются специфические переносчики, обеспечивающие транспорт ионов и некоторых соединений.
Так, в присутствии фосфатного транслокатора в хлоропласты поступает неорганический фосфат в обмен на триозофосфат. Дикарбоксилатный транслокатор обеспечивает поступление в строму органеллы и выход из нее дикарбоновых кислот, аденилатный осуществляет перенос через оболочку молекул АТФ. D.Laval-Martin предположил, что при становлении структуры хромопластов происходит изменение проницаемости мембран перистро-миума.
Изучая вопросы обеспечения стромы пластид предшественниками для синтеза терпеноидных соединений, M.Schneider и соавторы показали, что в процессе трансформации хлоропластов в хромопласты действительно изменяется проницаемость оболочки по отношению по крайней мере к некоторым соединениям. Авторы исследовали проницаемость мембран оболочки пластид на семи стадиях превращения хлоропластов в хромопласты.
Интактные хлоропласты не пропускали ни уксусной, ни мевалоновой, ни лимонной кислот в строму органелл. Однако в процессе превращения этих пластид в хромопласты M.Schneider и соавт. отмечали вначале появление, а затем резкое увеличение проницаемости оболочек к уксусной и мевалоновой кислотам, достигающее максимума, после чего она снижалась на последних этапах трансформации органелл.
При этом даже сформированные хромопласты, полностью лишенные хлорофилла, характеризовались довольно высоким уровнем проницаемости к мевалоновой и уксусной кислотам. На промежуточных этапах превращения также отмечалась проницаемость мембран оболочки для лимонной кислоты.
Выявленная способность мембран к проникновению мевалоновой и уксусной кислот появляется не одновременно, а максимальные значения проницаемости этих соединений достигаются на разных этапах развития хромопластов.
Специальными исследованиями, выполненными B.Liedvogel и H.Kleinig, было установлено, что хромопласты, подобно хлоропластам, сохраняют способность осуществлять с цитоплазмой обмен неорганического фосфата на триозофосфат с помощью фосфатного транслокатора. Эти же авторы показали, что в оболочке хромопластов функционирует также и аденилатный транслокатор, который может обеспечивать пластиду энергетическим потенциалом в виде АТФ, синтезируемой в цитоплазме клеток.
Перейдем к более подробному рассмотрении ультраструктуры хромопластов.
Внутрихромопластная мембранная система
Хлоропласты фотосинтезирующих тканей высших растений и водорослей содержат высокоразвитую мембранную систему в виде тилакоидов, организованных в граны.
В зависимости от исследуемого вида растений, физиологического состояния, стадии развития, условий питания и освещения, а также от источника пластид (листья, стебли и т.д.) наблюдаются изменения количества тилакоидов в гранах, числа гран на пластиду и некоторых других показателей состояния мембранной системы хлоропластов, однако наиболее общие характеристики внутримембранного комплекса (высокая упорядоченность, организация тилакоидов в граны) при этом остаются неизменными.
По сравнению с хлоропластами внутрипластидная мембранная система хромопластов развита намного слабее (за исключением хромопластов мембранного типа) и менее упорядочена. В то же время отличительной чертой хромопластов является разнообразие конфигурации мембранного комплекса. В.Т. Матиенко и ЕМ Чебану в монографии "Ультраструктура каротинои-допластов (хромопластов)" в качестве единицы организации мембранной системы хромопластов принимают тилакоид.
Этого же мнения придерживаются и подавляющее большинство других исследователей. Однако существует также другое мнение, что в хромопластах тилакоидов нет, хотя в них наблюдаются мембранные образования. Анализ указанных противоречий показал, что они обусловлены различной трактовкой термина "типа»-коид".
Б.Т.Матиенко и ЕМ.Чебану под этим термином понимают, как и было оригинально предложено W.Menke, замкнутое мембранное образование внутри пластид с непрерывным и внутренним пространством различной конфигурации (от диска до сильно извилистых тел) независимо от его состояния и степени развития.
Другие исследователи под термином "тилакоид" понимают организованные в граны хлорофилл-содержащие образования хлоропластов, функция которых связана главным образом с фотосинтезом, тогда как мембранные образования хромопластов они не именуют тилакоидами.
Следует отметить, что идентификация замкнутых участков мембран тилакоида, особенно у хромопластов со спирально закрученными мембранами, часто затруднена повреждениями в мембранах, вызванными плоскостью среза или недостаточно качественной подготовкой образцов для электронного микроскопирования. В то же время на некоторых снимках хромопластов удается обнаружить замкнутые окончания мембран даже для пластид с очень длинными тилакоидами.
Внутримембранная система хромопластов очень многообразна. Б.Т. Матиенко и Е.М.Чебану выделяют четыре основных состояния мембранных элементов хромопластов:
- парно-ламеллярное (каникулярное);
- везикулярное;
- концентрическое;
- трубчатое.
Авторы отмечают также, что, кроме основных конфигураций, в хромопластах находится множество промежуточных состояний, причем пластиды с промежуточными организациями мембранной системы встречаются намного, чаще, чем пластиды, которым присуща одна из основных конфигураций мембран. Это можно объяснить полиморфностью хромопластов во взрослом состоянии.
Кроме того, различные состояния мембранной системы могут отражать различные стадии онтогенетического становления как пластиды в целом, так и отдельных ее компонентов. По-видимому, некоторые конфигурации обусловливаются плохой сохранностью материала, следствием использования некачественных методик приготовления образцов для электронного микроскопирования или неверной трактовкой полученных результатов.
Мембранные образования, наблюдаемые в хромопластах различных растений как везикулы (пузырьки) могут быть разбросаны по всей пластиде или расположены в группах. Они различной величины и, видимо, различного происхождения и функционального назначения. Исследования, выполненные на хромопластах дыни в процессе ее созревания, а также на желтых и оранжево-красных пластидах других культур, показали, что пузырьки могут образовываться из:
- тилакоидов гран хлоропластов после их разбухания;
- отдельных участков межгранных ламелл, в результате локального вздувания и постепенного отделения;
- отростков инвагинаций внутренней мембраны оболочки.
В целом, как уже отмечалось, внутрипластидная мембранная система хромопластов развита значительно слабее по сравнению с хлоропластами фотосинтезирующих тканей, в которых мембранный комплекс занимает значительный объем. В то же время в хромопластах некоторых растений мембранная система развита очень интенсивно, составляя больше трех четвертей внутреннего объема органелл.
Наиболее ярким представителем таких хромопластов являются пластиды венца цветков нарцисса желтого (лженарцисса), на которых в последующем проводились интенсивные биохимические исследования. Внутрипластидная мембранная система этих органелл плотно прилегает к оболочке пластиды и состоит из нескольких очень длинных спирально закрученных тилакоидов, образующих до 20 слоев мембран.
Напротив, внутрипластидная мембранная система хромопластов волосков рыльца пестика кукурузы напоминает граны хлоропластов. Структурной основой мембран, в том числе и хромопластных, является липидный бислой в состоянии жидких кристаллов (или, как их еще называют, мезофаз) смектического типа.
Этот тип жидких кристаллов характеризуется двухмерной направленностью, при которой амфифильные липидные молекулы ориентированы перпендикулярно плоскости распространения бислоя (рис. 1). 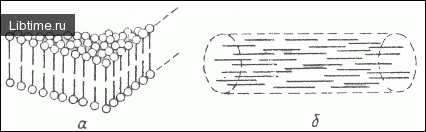
Основными липидными молекулами мембран хромопластов, как и других пластид, являются галактолипиды диацилмоногалактозилглицерин (ДАМГГ), диацилдигалактозилглииерин (ДАДГГ); серусодержащий гликолипид сульфохиновозилглицерин; а также фосфолипиды - фосфатидилглицерин, фосфатидилхолин. В мембранной фракции хромопластов содержатся также стерилгликозиды и их ацилированные молекулы.
В хлоропластах высших растений мембраны содержат также диацилтригалактозил-глицерин и диацилтетрагалактозилглицерин. В препаратах мембран хромопластов эти компоненты не обнаружены. Однако мембранные препараты хромопластов нарцисса желтого включали меченную УДФ-галактозу в диацил-тригалактозилглицерин, что предполагает существование в них галактолипидов с несколькими конденсированными остатками галактозы, хотя их количество, по-видимому, незначительно.
Препараты мембран хромопластов нарцисса желтого и настурции содержат около 5 % каротиноидов. В мембранах хромопластов перца однолетнего были обнаружены каротиноиды, а-токоферол. В препаратах мембран из пластид зрелых плодов перца не удалось обнаружить филлохинон (витамин Ki), тогда как хлоропластные мембраны содержали большое количество этого компонента.
Рассмотрим еще такой элемент ультраструктуры хромопластов, как белки.
Белки мембран хромопластов исследованы недостаточно. Известно, что часть из них представляет собой ферменты, участвующие в реакциях синтеза (гликозилировании и ацилировании) галактолипидов и стерилгликозидов, осуществляющие последние этапы образования а-токоферола, витамина Ki, а также превращающие первый синтезированный каротиноид (фитоин) в каротины и ксантофиллы, поскольку активности указанных ферментов выявлялись в мембранной фракции и отсутствовали в строме пластид.
При анализе пюлипептидного состава мембран хлоропластов нарцисса желтого в полиакриламидном геле после электрофореза в присутствии додецилсульфата натрия выявлено распределение белков, аналогичное образцам из тилакоидов других видов растений. Молекулярные массы преобладающих полипептидов находились в пределах 12-78 кД. Наиболее интенсивные полосы соответствовали полипептидам с молекулярными массами от 17 до 26, а также 52 и 58 кД.
Полипептиды внутрихромопластных мембран заметно отличались. Главное отличие заключалось в том, что в хромопластных образцах незначительны полипептиды с молекулярными массами ниже 30 кД. Большинство белковых компонентов мембран хромопластов характеризовались молекулярными массами от 50 до 150 кД с основными полосами в области 50, 58, 67 и 76 кД.
В то же время распределение по молекулярным массам полипептидов внутрипластидных мембран хромопластов отличалось от такового оболочки хлоропластов, хотя у последних также преобладали компоненты с высокими молекулярными массами. Следует отметить, что наблюдаемые различия между фракционным составом полипептидов мембран хромопластов и аналогичным составом оболочки хлоропластов носили больше количественный характер, чем качественный.
Многие полипептиды мембран хлоропластов были обнаружены в препаратах хромопластных мембран (о чем судили по электро-форетической подвижности), однако в измененных количествах. И только некоторые полосы, характерные для хлоропластных мембран, отсутствовали в хромопластных препаратах и наоборот. В то же время образцы распределения полипептидов хромо- и хлоропластных мембран практически не имели общих полос с белковыми компонентами митохондриальных мембран.
Сравнение полипептидов хромо- и хлоропластных мембран дало несколько неожиданные результаты. В частности, в составе хромопластных мембран были обнаружены полипептиды, принимающие участие в фотосинтезе. Это было продемонстрировано для большой и малой субъединиц рибулозобис-фосфаткарбоксилазы, субъединиц сопрягающего фактора фотофосфорилирования CFj, идентичность которых аутентичным полипептидам была подтверждена пептидным картированием.
Другие белки, принимающие участие в фотосинтезе, в препаратах мембран хромопластов не обнаружены. В частности, в них не были обнаружены компоненты, совпадающие по электрофоретической подвижности с полипептидами светособирающего комплекса II.
Сходная ситуация наблюдалась при исследовании белкового состава этиопластов, у которых также отсутствуют тилакоиды, не содержащие светособирающий комплекс II, в то время как у этих органелл были обнаружены субъединицы сопрягающего фактора фотофосфорилироваиия CFi и белки реакционного центра фотосистемы I.
На основании полученных результатов P.Hansmann, P.Sitte сделали предположение, что дифференциация отдельных типов пластид (хлоро-, хромопластов и др.) обусловлена не столько качественными изменениями в составе основных белковых компонентов, сколько накоплением отдельных видов белков, которые могут рассматриваться как "специфические" белки этого особого состояния пластидной дифференциации.
Подтверждающим материалом может быть то, что основные хромопластные белки, обозначенные авторами как Р4 и Р5, в хлоропластах представлены в следовых количествах. В отношении происхождения внутрипластидной мембранной системы хромопластов существуют две точки зрения. Большинство исследователей придерживаются первой из них, согласно которой мембранная система хромопластов образуется в результате впячивания и последующего роста внутренней мембраны оболочки органеллы.
Однако P.Gronegress высказал предположение, что внутрипластидные мембраны хромопластов, как и другие пигментнесущие структуры, развиваются из тилакоидов гран хлоропластов.
Структурные данные о развитии хромопластов нарцисса желтого, а также результаты биохимических исследований, находятся в противоречии с предположением P.Gronegress и склонны объяснить возникновение внутрипластидных мембран хромопластов как результат впячивания, роста и развития внутренней мембраны оболочки органеллы.
В частности, внутрипластидные мембраны и оболочки хромопластов характеризовались низким содержанием белка и пониженной плавучей плотностью.гладкой поверхностью после замораживания-оттаивания и негативного окрашивания препаратов, имели сходный состав липидов и высокое соотношение ДАДГГ к ДАМГГ.


