Каротиноиды
Как отмечалось ранее, хромопласты характеризуются способностью накапливать большие количества каротиноидов. Каротиноиды представляют собой соединения терпеноидной природы и большинство из них принадлежат к тетратспенам, содержащим 40 углеводных атомов в молекуле (С40 -соединения).
Они состоят из восьми изопреновых единиц и образованы связыванием "хвост к хвосту" двух фрагментов, каждый из которых состоит из четырех изопреновых остатков, соединенных "голова к голове". Таким образом, две центральные метильные группы находятся в 1,6-положении относительно друг друга, в то время как остальные нетерминальные метильные группы находятся в 1,5-положении (рис. 1). 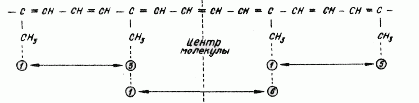
Каротиноиды.Общая характеристика
Все каротиноиды формально могут быть получены из ациклического соединения ликопина (рис. 2) посредством реакций, включающих гидрогенирование, дегидрогенирование, циклизацию, вставку кислорода в различные положения, миграцию двойных связей, миграцию метальных групп, удлинение цепи, укорочение цепи. 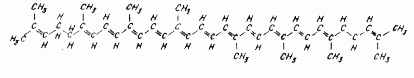
Каротиноиды, состоящие исключительно из атомов углерода и водорода, называются каротинами. К ним относятся ликопин, фитоин, фитофлуин, 'alpha;, 'beta;, 'gamma;, 'delta;, 'zeta;, 'epsilon;-каротины, нейроспорин, 'alpha;- и 'beta;-зеакаротины (рис. 4). Каротиноиды, содержащие кислород, называются ксантофиллами. Подавляющее большинство известных в настоящее время каротиноидов - ксантофиллы (рис. 4).
Каротиноиды, у которых одинарные и двойные связи смещены на одну позицию, называют ретрокаротиноидами. К ретрокаротиноидам, например, относится пигмент ксантофильной группы эшшольцксантин. 
Кроме С40 -каротиноидов в растениях распространены их производные, которые содержат меньше 40 атомов углерода (апокаротиноиды), примером которых могут служить 3-цитраурин и кроцетин. У грибов и бактерий встречаются также С45 - и С50 -каротиноиды, не обнаруженные у высших растений. Наличие сопряженных двойных связей в структуре каротиноидов может обусловливать цис-транс-томерно.
Большинство природно встречающихся каротиноидов находятся в транс-форме. Однако у живых организмов, в том числе и у растений, обнаружены также цис-изомеры некоторых каротиноидов, например цис-фитоин, цис-фитофлуин, проликопин (цис-изомер ликопина). Циклические структуры во многих каротиноидах содержат асимметрические атомы углерода, что также обусловливает существование множества стереоизомеров.
В частности, хризантемаксантин и флавоксантин имеют одинаковую структурную формулу, но различаются между собой пространственной ориентацией боковых группировок. 
Каротиноиды встречаются в свободном состоянии или могут быть этерифицированы жирными кислотами, ацетатом и углеводами.
Сложные эфиры ксантофиллов с пальмитиновой, стеариновой, миристановой, лауриновой кислотами и ацетатом обнаружены в лепестках цветков подсолнечника однолетнего, а основное количество кроцетина, наиболее обильно представленного пигмента лепестков сафрона, этерифицирова но гентиобиозой и глюкозой в различных сочетаниях.
Распространение и локализация каротиноидов
Каротиноиды фотосинтезирующих тканей локализованы в основном в гранах хлоропластов, вероятно, в форме хромопротеидов. В частности, были обнаружены комплексы белков с виолаксантином и 'beta;-каротином. Когда хлоропластные белки солюбилизируются детергентом, они могут быть разделены с помощью центрифугирования на две основные фракции - легкую и тяжелую, которые соответствуют фотосистемам I и II.
Каротиноиды неравномерно распределены между этими двумя фракциями. Фотосистема I обогащена 'beta;-каротином, в фотосистеме II преобладают ксантофиллы. Пигменты этиолированных проростков локализованы в этиопластах. При этом следует отметить, что преобладающие пигменты в этиопластах этиолированных проростков и хлоропластов зрелых листьев отличны между собой.
Так, основными ксантофиллами этиопластов фасоли обыкновенной являются флавоксантин и хризантемаксантин, которые отсутствуют в зеленых листьях. В то же время в них не обнаруживается неоксантин, который является наиболее обильным пигментом в листьях взрослых растении. Каротиноиды в лепестках цветков локализованы в хромопластах.
В хромопластах желтого нарцисса каротиноиды накапливаются в основном в многочисленных концентрических мембранах. 'beta;-Каротин в пластидах венца нарцисса снежно-белого находится в кристаллах, расположенных во внутритилакоидном пространстве. В хромопластах цветков хризантемы посевной и испанского дрока обыкновенного, тюльпана, саротамнуса метлистого и многих других растений каротиноиды локализованы в осмиофильных пластоглобулах.
В лепестках калюжницы болотной каротиноиды, помимо хромопластов, обнаруживаются также в хлоропластах, а в цветках некоторых растений каротиноиды отсутствуют.
Ксантофиллы в хромопластах цветков, в отличие от пигментов фото-синтезирующих тканей, этерифицированы пальмитиновой, стеариновой, миристиновой либо лауриновой кислотами. Обнаружены также каротиноиды, этерифицированные ацетатом и углеводами. Зрелые плоды многих растений окрашены благодаря наличию в них тех или иных каротиноидов.
Как и в цветках, каротиноиды плодов локализованы в хромопластах, которые развиваются из хлоропластов в процессе созревания. В некоторых случаях, как например в плодах ландыша майского, хромопласты образуются из пропластид.
Каротиноиды в хромопластах красных плодов перца однолетнего, тыквы обыкновенной, розы морщинистой и плодах некоторых других растений локализованы в осмиофильных пластоглобулах и трубчатых образованиях. В плодах желтых, оранжевых и белых разновидностей перца однолетнего каротиноиды накапливаются в форме кристаллических образований.
Ксантофиллы в плодах, как и в цветках, в значительной степени этерифицированы. Распространены каротиноиды в подземных органах моркови и батата, хотя следует отметить, что цвет некоторых азиатских разновидностей моркови обусловлен наличием антоцианов. 90-95 % каротиноидов оранжевых сортов моркови представлены каротинами.
Среди них наиболее обильно представлены 'alpha; 'beta;, v-каротины и ликопин, в то время как 'gamma;-каротин, 'zeta;-каротин, нейроспорин, фитоин и фитофлуин обнаружены в следовых количествах. Ксантофиллы в оранжевой моркови составляют только 5-10 % общего количества каротиноидов, однако их количество возрастает до 75-93 % в разновидностях желтой моркови и не менее 95 % в белой моркови.
Основным пигментом батата (Ipomea batatas edulis) является 'beta;-каротин. В моркови пигменты локализованы в хромопластах кристаллического типа, структура которых была детально изучена. Каротиноиды также обнаружены в семенах, пыльниках, тычинках, пыльце различных растений. Показано, что в придатках початков тифониума расщепленного и арума они локализованы в хромопластах. Каротиноидный состав хромопластов весьма своеобразен и существен но отличается от состава пигментов в хлоропластах.
Несмотря на то, что основные каротиноиды большинства хромопластов обнаружены также и в хлоропластах фотосинтезирующих тканей, их количественное соотношение в этих органеллах различное. В то же время в хромопластах некоторых растений находятся специфические каротиноиды, которых нет в хлоропластах. Так, например, капсантин - один из преобладающих пигментов зрелых томатов - содержится только в хромопластах.
Более того, это видоспецифический пигмент, поскольку его до настоящего времени не удалось обнаружить у других растений. Как отмечалось ранее, основная масса каротиноидов растений локализована в пластидах. Однако каротиноиды были идентифицированы и в непластидных структурных компонентах растительных клеток.
В частности, многие зеленые водоросли при неблагоприятных условиях развития, обычно при азотном голодании, накапливают большие количества каротиноидов во внутриклеточных отложениях без ограничивающих мембран и в липидных вакуолях. S.Brown и J.Prebble, применяя специальные предосторожности с целью ингибирования липаз и полифенолокскдаз, обнаружили, что распределение каротина во фракциях при дифференциальном центри-фугировании в градиенте плотности сахарозы гомогената цветной капусты совпадало с распределением сукцинатдегидрогеназы - фермента, являющегося маркером для митохондрий.
На основании этих экспериментов авторы заключили, что митохондрии содержат каротиноиды. Аналогичные выводы сделаны в опытах с клубнями картофеля, где каротиноиды обнаружены также в других фракциях, в частности во фракции "легких" мембран и в микросомах. Однако количество пигментов в непластидных фракциях было незначительным, что несколько затрудняет интерпретацию полученных результатов.
Каротиноиды обнаружены также у грибов, бактерий и животных. Показано,что в отличие от высших растений, водорослей, бактерий и грибов, животные сами не могут синтезировать эти соединения, которые поступают в их организм вместе с пищей, после чего модифицируются в специфические "животные каротиноиды" особыми ферментными системами.


