Метаболизм углеводов
Активный синтез и накапливание липидных соединений, а также каротиноидов требуют постоянного обновления пула предшественников источников энергии и восстановительных эквивалентов в виде НАДН2 и НАДФН2.
Хлоропласта фотосинтезирующих тканей обеспечиваются этими компонентами благодаря фотосинтетическим реакциям. В отличие от хлоропластов хромопласты - метаболически гетеротрофные органеллы.
Опыты показали, что активные в синтезе липидов и каротиноидов изолированные хромопласты практически не осуществляют свето-зависимого включения углекислого газа. Поэтому обновление пула органических соединений - предшественников синтеза каротиноидов и липидов, а также источников энергии и восстановительных эквивалентов этих органелл не может обеспечиваться посредством фотосинтеза.
Рассмотрим подробнее процесс метаболизма углеводов.
H.Ziegler и соавт. показали, что в процессе превращения хлоропластов плодов однолетнего перца в хромопласты, помимо потери свето-зависимой способности фиксировать углекислый газ, происходят также существенные изменения в активности различных ферментов. углеводного обмена.
Показано, что первичные этапы фиксации углекислого газа, которые приводят к образованию глицерин-3-фосфата и яблочной кислоты, в процессе трансформации пластид меньше блокируются, чем последующие реакции. В то же время пластиды с промежуточной организацией структуры (хлорохромопласты) сохраняли довольно активные ферменты, осуществляющие превращение первичных продуктов фиксации углекислого газа в глюкозо-6-фосфат- субстрат для окислительного пентозо-фосфатного цикла превращений углеводов. 
Выяснилось, что характеристические ферменты цикла Кальвина (рибулозобисфосфат-карбоксилаза, фосфорибулокиназа, НАДФ+ -зависимая глицеральдегидфосфат-дегидрогеназа) теряют в процессе трансформации около 80% активности, тогда как фруктозо-бисфосфат альдолазная активность не изменялась.
Авторы объясняют высокую альдолазную активность в хромопластах высокой эффективностью использования гексозофосфатов в качестве субстратов для окислительного пентозо-фосфатного цикла.
Подтверждением этому служили эксперименты, показывающие, что ключевой фермент этой цепи превращений углеводов глюкозо-6-фосфат дегидрогеназа имеет сходные высокие активности в хлоро- и хромопластах, а в органеллах с промежуточной организацией они были в 1,4-1,5 раза выше.
Известно, что в фотосинтетически активных хлоропластах активность глюкозо-6-фосфат дегидрогеназы фоторегулируется. В зрелых хромопластах это не происходит. В качестве основного субстрата для синтеза липидов и каротиноидов хромопласты используют ацетил-КоА. Последний может образовываться из уксусной кислоты под действием фермента ацетил-КоА синтетазы, которая была обнаружена в функционально активных пластидах желтого нарцисса.
Опыты с пластидами in vitro показали, что в процессе трансформации хлоропластов в хромопласты изменяется проницаемость мембран оболочек пластид к некоторым соединениям, в том числе и уксусной кислоте (см. Оболочка хромопластов). Ацетил-КоА (а также АГФ и НАДН2) могут образовываться в результате гликолитических превращений из триозофосфатов.
Результаты исследований H.Kleinig, B.Liedvogei показали, что глицерин-3-фосфат и пировиноградная кислота могут служить предшественниками жирных кислот в хромопластах желтого нарцисса. Очевидно, в хромопластах осуществляется цепочка гликолитических превращений, ведущих от диоксиацетонфосфата до ацетил-КоА.
Триозофосфаты могут также поступать в хромопласта из цитоплазмы в обмен на неорганический фосфат через фосфатный транслокатор в хромопластной оболочке, где функционирует также аденилатный транслокатор, который может обеспечивать эти органеллы АТФ, синтезированной в цитоплазме.
В синтезе жирных кислот и каротиноидов в качестве кофермента на отдельных этапах биохимических превращений используется НАДФН2. По-видимому, НАДФН2 синтезируется в хромопластах с помощью реакций окислительного пентозо-фосфатного цикла.
Обнаруженные высокие альдолазная и глюкозо-6-фосфат дегидрогеназная активности в хромопластах являются аргументами в пользу такого предположения. Метаболизм углеводов лучше представить в виде изображения. 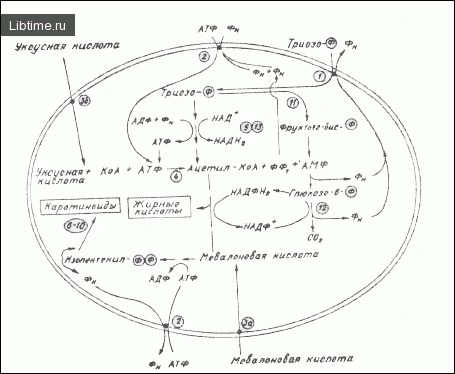
1 - фосфатный транслокатор, 2 - аденилатный транслокатор, 3 - проникновение уксусной и мевалоновой кислот через мембраны-оболочки хлоро-хромопластов и хромопластов, 4 - ацетил-КоА синтетаза, 5-- гликолиз, 6-70 - биосинтез каротиноидов из изопентенилгарофосфата, 11 - альдолаза, 12 - глюкозо-6-фосфат дегидрагеназа, 13 - НАД+ -зависимая триозофосфат дегидрогеназа.
На основании этих данных H.Ziegler с соавт. предложили схему наиболее важных метаболических процессов в хромопластах (рис. 1). H.Kieinig, B.Liedvogei исследовали эффективность различных экзогенных источников энергии в синтезе жирных кислот. Помимо АТФ, хромопласты венца лженарцисса относительно эффективно использовали АДФ в качестве источника энергии (74 % включения [1-14С] -уксусной кислоты в жирные кислоты по отношению к АТФ).
H.Kleinig, B.Liedvogel считают, что высокая эффективность АДФ обусловлена функционированием в хромопластах аденилаткиназы, присутствие которой ранее было показано в фото-синтезирующих органеллах различных организмов. УТФ, ЦТФ и ГТФ оказывали слабый стимулирующий эффект (15-32 % по сравнению с АТФ), а АМФ практически не стимулировал включения метки в жирные кислоты.
Известно, что гликолитические реакции, ведущие от диоксиацетонфосфата до ацетил-КоА, в качестве промежуточных соединений используют высокоэнергетические органические молекулы. H.Kleinig, B.Liedvogel испытали высокоэнергетические соединения - промежуточные вещества в цепи гликолитических превращений - в качестве возможных источников энергии.
Диоксиацетонфосфат, глицеральдегид-3-фосфат и глицерин-3-фосфат характеризовались активностью, сравнимой с АТФ. В то же время глицерин-2-фосфат и фосфоенолпировиноградная кислота эффективнее стимулировали синтез жирных кислот (в 2,3-2,7 раза) по сравнению с АТФ.
Было показано, что хромопласты содержат пируваткипазу, которая катализирует реакцию образования АТФ из фосфоенолпирувата.Изолированные хромопласты содержали всего 50 пМ АТФ/мг белка, что соответствует 5 мкМ. В то же время после добавления к хромопластной суспензии фосфоенолпирувата количество АТФ в хромопластах увеличивалось в 7-8 раз благодаря фосфорилированию эндогенного АДФ или АМФ и составило 380 пМ АТФ/мг белка, или 36 мкМ.
Таким образом, было установлено, что промежуточные продукты окислительного пентозо-фосфатного цикла могут быть эффективными источниками энергии для биосинтеза липидов и каротиноидов. Если это имеет место в хромопластах in vivo, то не исключено, что фосфатный транслокатор в оболочке хромопластов осуществляет обратную реакцию, т.е. обеспечивает поступление триозофосфатов в строму пластид в обмен на неорганический фосфат.
Отмечалась оперативность обратной функции фосфатного транслокатора в хлоропластах при нефотосинтезирующих условиях.


